

eye see, therefore i am

“The Eye of Nature” by Dan Hilliard
Seen firsthand, there is only one thing in the universe, if at all… and it is the eye itself. And even the thingness of that ‘thing’ is behind the lens.
Sight is not out there; it is in here.
Consider the monadic, enclosed, singly-lensed eye as a concave theater, the fovea of the retina the stage, and the retina itself a wet and less differentiated compound eye, in contrast to the convex and discretized eye of the insect. The architecture of the monadic eye converges perception toward focal point, as opposed to the diffuse and highly spatially complex field of the insect. This contrast tectonically steers the entire informational and symbolic order downstream of lensed sight.
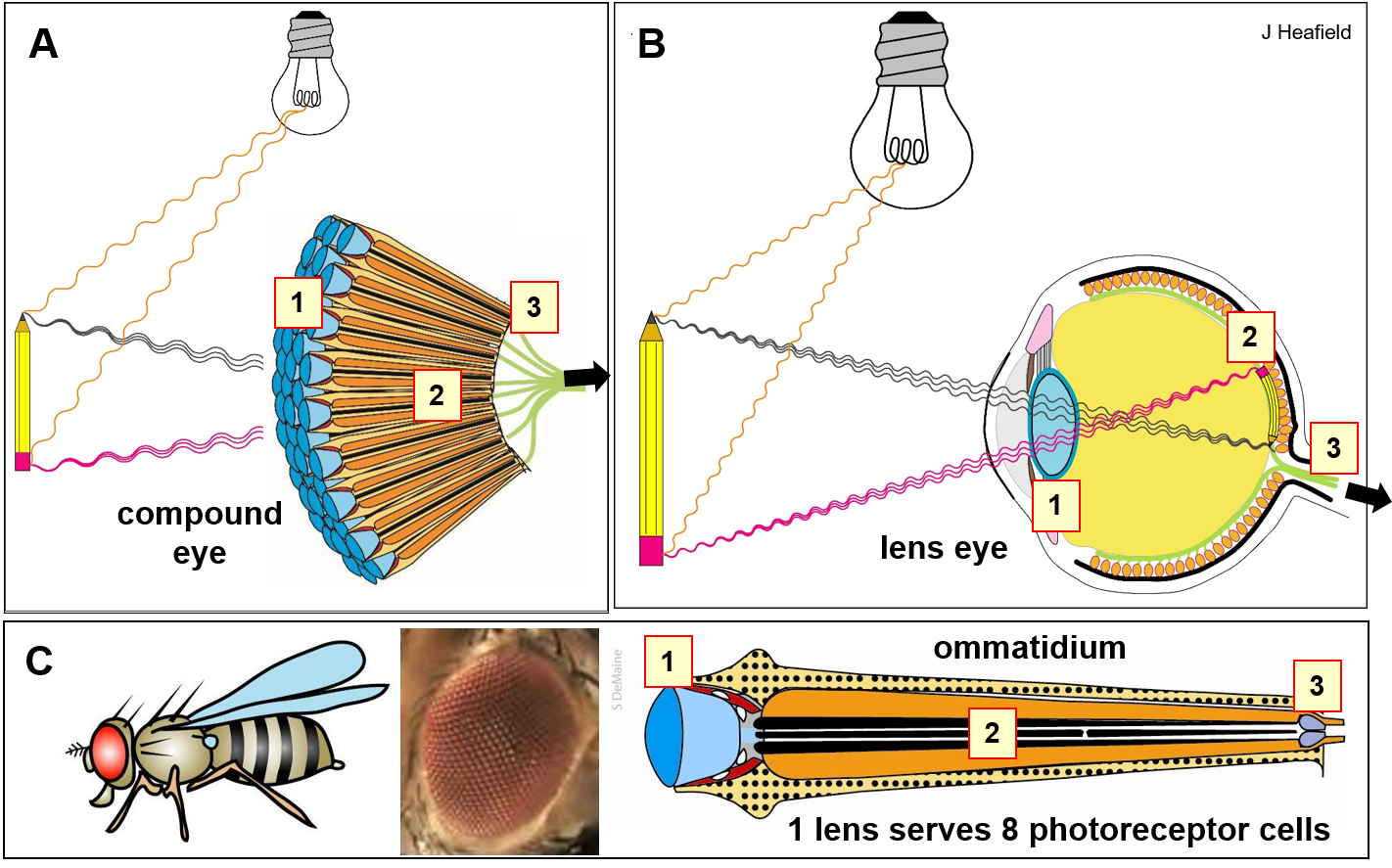
From this comparison alone, it is reasonable to conclude that the abstraction of biological vision emerges in metabolic choreography and protein goo in its primacy, and then further into higher dimensional compound abstraction in the anatomy and optics of a given eye atop the photoreception.
Seeing as sight is fleshy, the notion of the Cartesian theater crumbles! That which would hold that the eye is beaming onto a fully disembodied projector screen somewhere in the aether, somehow also phenomenologically behind the eyes, cannot hold its own assertion up.
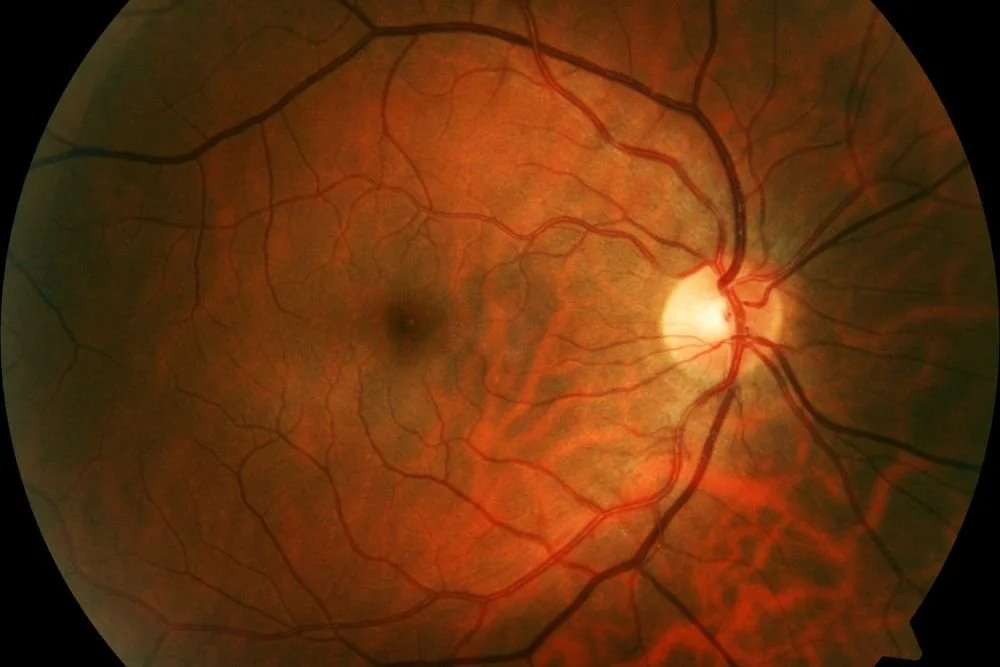
Retinal vasculature; the shadowy spot within the fine branches is the fovea, the bright locus is the optic nerve.
The blood vessels enabling this all in our eyes are, after all, positioned in front of the retina, pulsing away, constantly being adjusted for both their motion and the reticulate branching negative space they form, more unique than a fingerprint.
Further on this uniqueness, no photoreceptor’s specific reception of light, no ocular lens shape, no ocular atrium geometry, no vitreous humor, no retinal metabolism nor metabolic byproduct clearance nor vasculature nor vascular efficiency nor rod/cone-nerve junction anatomy, nor optic nerve organization nor optic nerve metabolism, nor nor nor nor nor… not one of the uncountable single links in the vast strange loop chain of vision are the same across single bodies, even across the midline of a body from one eye to the other.
So not only is it fleshy, it is specific to our flesh, your seeing yours and my seeing mine. Each of us see through the shadow of the body, the ripples of our heartbeats. Our sight, in so many ways, rumbles and glows likewise with the inmost choreography of our very living itself.
The body is then empirically in the way of the “perfect” image—yet there is no image without body.
The body is choreographically and architecturally organized around maintaining, participating in, and processing vision.
It is well known that an outsized proportion of metabolic activity in the body occurs in the small locus of the brain, and that the majority of this metabolic activity goes to integrating vision with somatic and sensory information to construct the controlled synesthesia that steers perception into a form woven with motor planning that is a substrate for phenomenological consciousness.
But the most metabolically intensive issue per cell is not the brain—it is the retina.
Retinal tissue is the most metabolically active tissue in the entire body, consuming 50% more oxygen and producing orders of magnitude more of numerous metabolic byproducts than brain tissue.
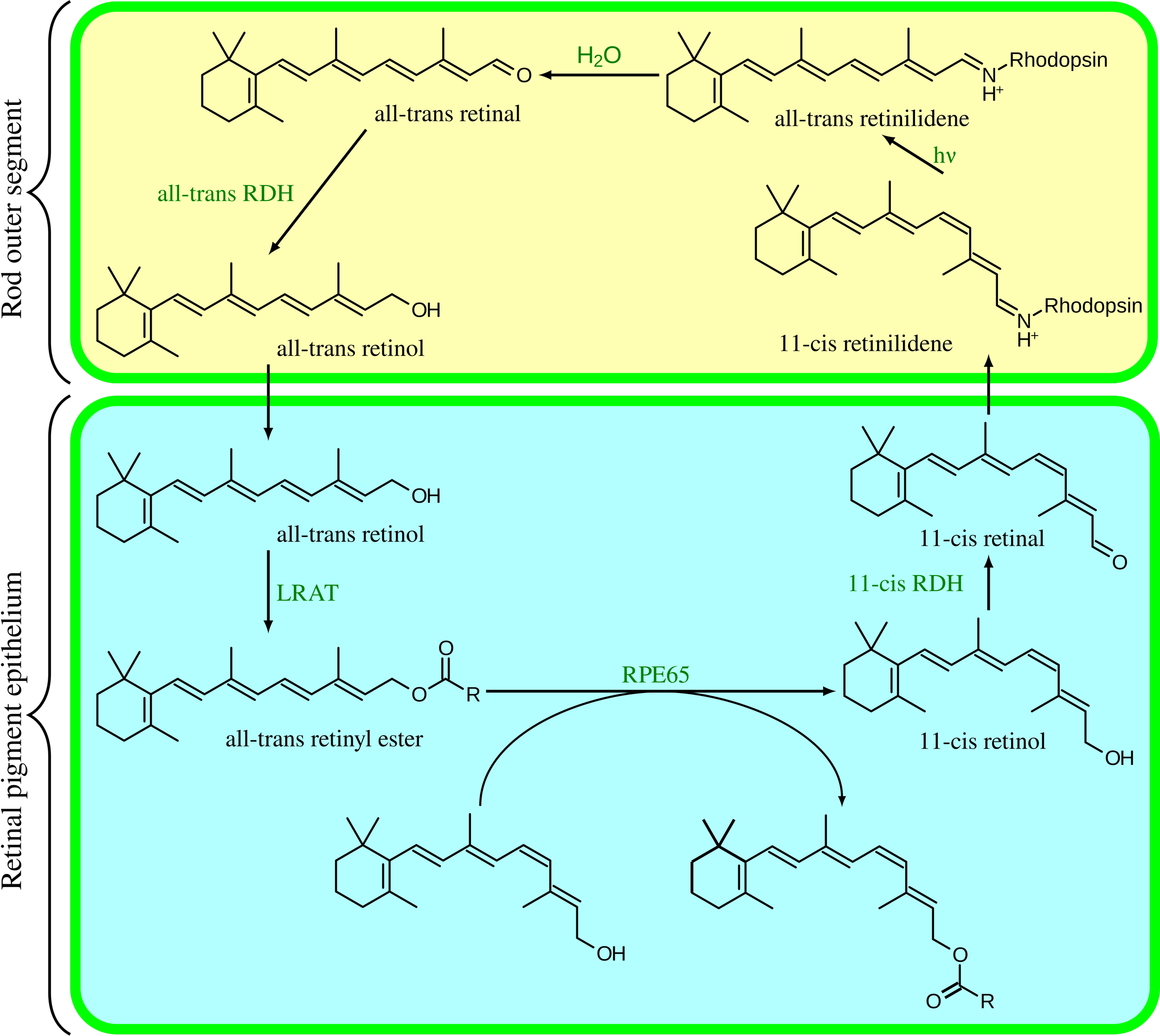
The visual cycle, one among many processes renewing constantly at high volume in the retina. Very metabolically expensive.
Doubling down on this, the eye’s rectus muscles contract more frequently in a day than even the heart. Heart, the diaphragm, the digestive system, and the musculoskeletal ensemble—of all muscles in the entire body, it is those of eye that are most frequently activated.
Our ways of seeing are part and parcel of our choreographies of living, moving too—seeing as the dance of the eyes is our primary motor focus.
Therefore there is no platonic eye. All the eyes that exist and ever have still do not saturate the space of all possible eyes.
It is also quite difficult to delineate the eye from brain; the retina and optic nerve can be thought of as extensions of brain matter out of the skull, though evolutionarily this development went in reverse.
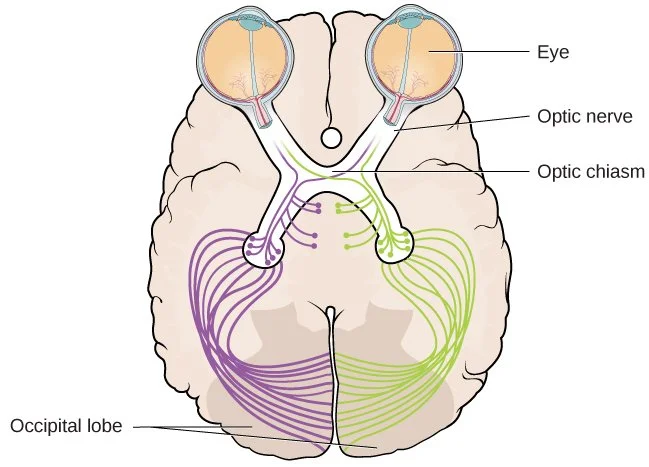
The eyes are the terminus of a dense tract of brain tissue protruding out of the skull by way of the eye sockets.
This genealogy of eye before brain echoes in endogenous psychopharmacology. Serotonin receptors, among many other G-protein coupled receptors integral to the coordination of brain activity and inference across sensory and somatic inputs to help maintain the organs, are rhodopsin-like proteins, evolutionarily derivative of the opsin proteins that respond to light in photoreceptive tissues across the ecological web, from the human retina down to the dinoflagellate erythropsidinium, which has the smallest codified eye structure known as part of its single cell.

Model of a generic rhodopsin receptor protein crystal structure.
Also subtextual here is that the rhodopsin receptor family are a subtype of the opsins—which is to say, opsins came first. This is supported in the evolutionary tree, in fact, our friend erythropsidinium can help us. Possessing an eye as a single cell, lacking distinct neural tissue, there is a direct evolutionary referent demonstrating the utilitarian primacy of the eye, and the optically oriented nervous system as a secondary force multiplying faculty.
Not only is eye the progenitor of brain, it is the progenitor of the architecture of multimodal somatosensory entanglement in the mammal and other organisms with nervous systems. The neural faculties of sight are not, theoretically, constrained from making sense of that beyond sight, or adjacent to sight; in fact, this is how sensory cognition works. The emergent cymatic of sensory fields perturbing form an informational map for the cortex to inference on and engage, or refrain from engaging, motor pathways. But I digress on this.

erythropsidinium
Back to the earlier point: If erythropsidinium sees with an eye in lineage with ours, we must consider what that means for the place of our sight in the overall family of seeing beings. Is our sight the conclusive sight? Preposterous. Is single focal point sight the conclusive way of seeing for life? Preposterous, too.
This evident arbitrariness of sorts leads to an interesting juncture.
Seeing as our opsins can only register signals from the wavelengths that pass through water in the vitreous body within the eye (the rainbow)—an artifact from our marine evolutionary ancestors’ ways of life—there are already very specific incentives baked into our psychophysics.
Let’s extend the chain of contingency—seeing through a liquid medium behind a liquid-filled lens, and then through air, there is already an inherent abstraction built into the optics alone. Absent the ampullae of Lorenzini possessed by sharks to detect prey electrically at range, or their lateral line which senses vibration at distance (elegantly collapsing the dichotomy between sound and touch, by the way,) evolution converged on ranged optical perception as the primary mode of environmental externally focused perception and vigilance for us, so we are in a class still narrower within all of ecology.
Yet further, we see this spectrum in a spatial mode oriented toward single point focus at distance, collapsing the visual field into a flattened spatial plane, three dimensional inference arising only from calculating divergence between two flat planes; the spatial world is thus a well informed daydream. And then in signal processing, from the retina to the brain, rapid (high frequency) difference is the primary cue for perceptual registry, whether from luminant or chromatic contrast, or from the motion of an object at some distance, or the movement of our own body in locomotion or active stabilization while standing reveals parallax and thus detail.
What doesn’t move, or what is extremely slow, goes essentially unnoticed except if we move in reference to it.
Seeing anything, anything at all, as we do, is incredibly contingent on the precise and very specific situation the human being finds themselves in within the spatial environment and incentive structures of survival.
If the architecture of our sight and seeing is this narrowly situated, we have a lot less ground to stand on regarding any semblance of knowing with conclusion even from sight, perhaps the most informationally salient sense (salience vis a vis language, conceptual thinking, situationally reactive thought; touch, proprioception, and subperceptual interoception are the bedrock of all somatosensory inference).
And so firsthand, it simply must be that there is no “perfect” image, no true image, not even to the eye itself; the eye may be the very barrier between image and truth altogether. Sight is not unbroken, sight is not final; sight is transitory, vision is passage.
The glances never contained the whole picture. The fragments never fit perfectly, and yet there appears a wholeness in their assemblage either way.
Sight is not without filter. Sight, more accurately, is what survives after available information in the form of energy passes through an extremely fine and contingent stack of filters.
The basis of the image is not what’s there; the basis of the image is the removal of essentially all that is.
So what even is an eye? Is “the eye” even confined to the surface-facing anatomical structure? Seeing as the localized brain evolved after the localized eye, a strong argument could be made that it is not confined whatsoever to the ocular atrium. Far from it. The ocular atrium isn’t even necessary.
The eye is, in fleshy terms, secondary to anatomical void, built upon with tissue through which light enters and affects an organism. The eye is a sort of high dimensional pore, or even a benign, maybe sinthomic, congenital wound—a reverse symptom buffering the harsh existential conditions impinging on the organism, a point where contradictions meet and hold one another steady against collapse—quite literally a wound where the light enters.
Perhaps, more simply put, the eye is void rupturing void, form within emptiness, emptiness within form.

being and nothingness, 2024